
Complexity Thoughts: Issue #70
Unraveling complexity: building knowledge, one paper at a time
If you find value in #ComplexityThoughts, consider helping it grow by subscribing and sharing it with friends, colleagues or on social media. Your support makes a real difference.
→ Don’t miss the podcast version of this post: click on “Spotify/Apple Podcast” above!
Foundations of network science and complex systems
Evolutionary games with environmental feedbacks
Game theory usually brings to mind rivalries at the poker table or in business, but in reality, strategic interactions shape nearly every living system, from microbes to humans. What makes things especially fascinating is that these strategies don’t play out in a vacuum: they constantly reshape the very environment in which they unfold, which then loops back to change the “rules of the game”.
Farmers overharvest a resource, fishers deplete a stock or plants alter soil nutrients: each action shifts the environment, which in turn shifts the incentives for future actions. This paper proposes a unifying framework called eco-evolutionary game theory, designed to capture exactly this dance between behavior and environment. By modeling ecosystems that grow, decay or tip into new states, the authors reveal how outcomes hinge on whether individuals are quick to adapt or slow to follow, and on whether environments change faster or slower than strategies.
This framework applies to any game-theoretic interaction where the strategies that individuals employ impact the environment through time, and the state of the environment conversely influences the payoffs of the game. Such feedbacks are common in social-ecological systems, evolutionary ecology, and psychology.
What emerges is a set of patterns that cut across disciplines, offering insights into everything from collective decision-making to sustainable harvesting, and reminding us that strategy and environment are never separable: they coevolve.
This paper would naturally fit the “Evolution” section of the blog, but its results are so general that I find it more comfortable to consider it as foundational.
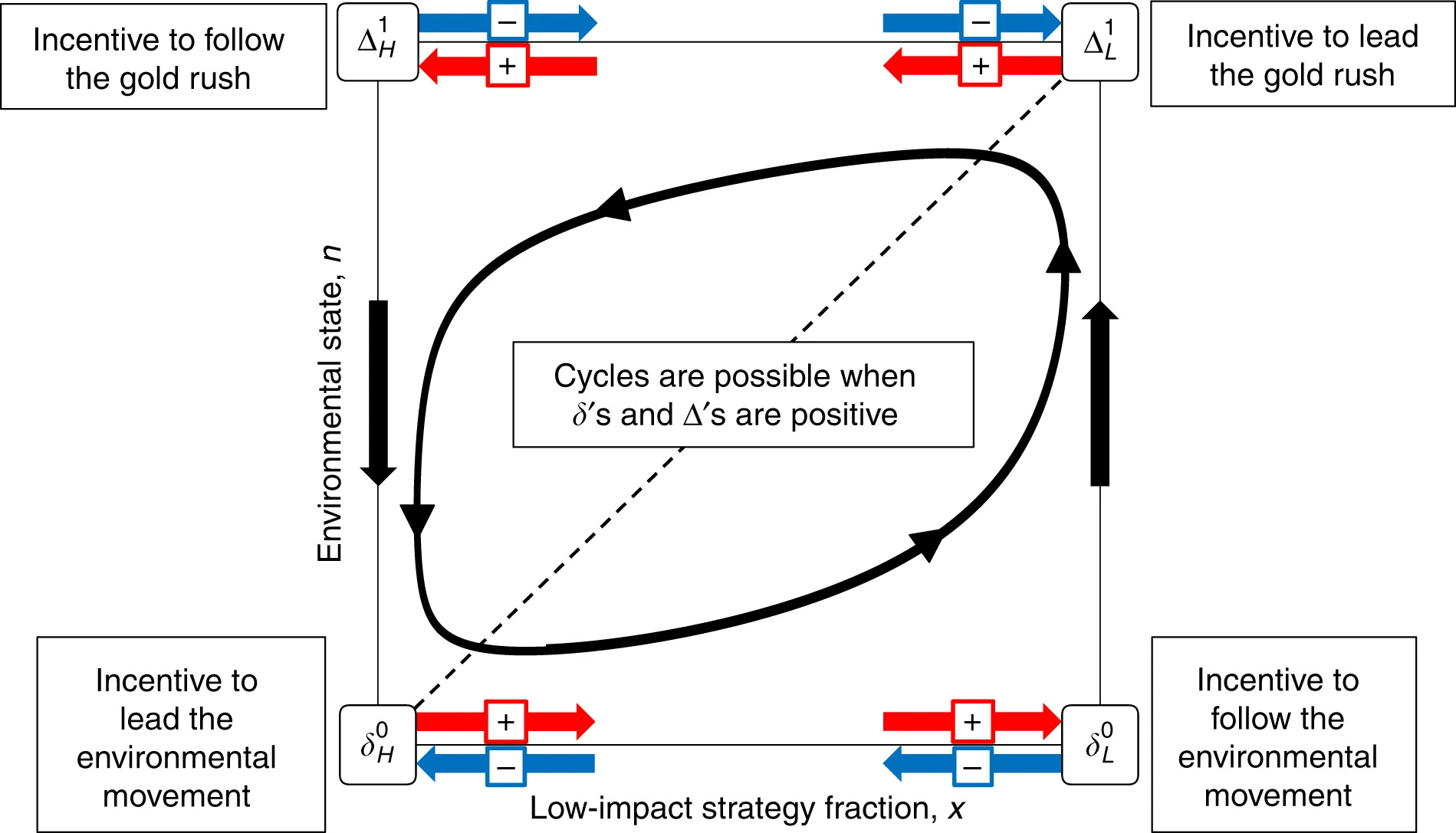
Strategic interactions arise in all domains of life. This form of competition often plays out in dynamically changing environments. The strategies employed in a population may alter the state of the environment, which may in turn feedback to change the incentive structure of strategic interactions. Feedbacks between strategies and the environment are common in social-ecological systems, evolutionary-ecological systems, and even psychological-economic systems. Here we develop a framework of ‘eco-evolutionary game theory’ that enables the study of strategic and environmental dynamics with feedbacks. We consider environments governed either by intrinsic growth, decay, or tipping points. We show how the joint dynamics of strategies and the environment depend on the incentives for individuals to lead or follow behavioral changes, and on the relative speed of environmental versus strategic change. Our analysis unites dynamical phenomena that occur in settings as diverse as human decision-making, plant nutrient acquisition, and resource harvesting. We discuss implications in fields ranging from ecology to economics.
Ecology & Evolution
Competition for resources can reshape the evolutionary properties of spatial structure
Many evolving ecosystems have spatial structures that can be conceptualized as networks, with nodes representing individuals or homogeneous subpopulations and links the patterns of spread between them. Prior models of evolution on networks do not take ecological niche differences and eco-evolutionary interplay into account. Here, we combine a resource competition model with evolutionary graph theory to study how heterogeneous topological structure shapes evolutionary dynamics under global frequency-dependent ecological interactions. We find that the addition of ecological competition for resources can produce a reversal of roles between amplifier and suppressor networks for deleterious mutants entering the population. We show that this effect is a nonlinear function of ecological niche overlap and discuss intuition for the observed dynamics using simulations and analytical approximations. We use these theoretical results together with spatial representations from imaging data to show that, for ductal carcinoma, where tumor growth is highly spatially constrained, with cells confined to a tree-like network of ducts, the topological structure can lead to higher rates of deleterious mutant hitchhiking with metabolic driver mutations, compared to tumors characterized by different spatial topologies.
Ecological Specialization and Trade Affect the Outcome of Negotiations in Mutualism
By definition, mutualisms involve the exchange of goods or services between partners. It has been shown that mutualism can grade into parasitism, but even when exchange is mutually beneficial, a conflict of interest remains because each partner benefits from reaping more benefits at a lower cost. Metaphorically, the partners negotiate the conditions of trade, the outcome of which will determine the net benefit to each partner. Each partner can adjust its allocation to self-provisioning while negotiating the ratio at which benefits are exchanged. To understand how these two features of trade affect mutualisms, we used the example of the plant–arbuscular mycorrhizal mutualism and modeled uptake and trade of two resources, phosphorus and carbon. In most contexts, the fungus specialized on phosphorus uptake while the plant took up both phosphorus and carbon. However, when phosphorus was abundant and light was scarce, the plant specialized, taking up only carbon and relying on trade for phosphorus. Resource availability was the most important factor determining specialization and the outcome of negotiation and trade, but other aspects of the context were also important. These results suggest experiments to link these two key features of trade with environmental conditions to determine the outcome of mutualism.
Social-ecological systems as complex adaptive systems: modeling and policy implications
Systems linking people and nature, known as social-ecological systems, are increasingly understood as complex adaptive systems. Essential features of these complex adaptive systems – such as nonlinear feedbacks, strategic interactions, individual and spatial heterogeneity, and varying time scales – pose substantial challenges for modeling. However, ignoring these characteristics can distort our picture of how these systems work, causing policies to be less effective or even counterproductive. In this paper we present recent developments in modeling social-ecological systems, illustrate some of these challenges with examples related to coral reefs and grasslands, and identify the implications for economic and policy analysis.
Social polymorphism is favoured by the co-evolution of dispersal with social behaviour
Dispersal determines gene flow among groups in a population and so plays a major role in many ecological and evolutionary processes. As gene flow shapes kin structure, dispersal is important to the evolution of social behaviours that influence reproduction within groups. Conversely, dispersal depends on kin structure and social behaviour. Dispersal and social behaviour therefore co-evolve, but the nature and consequences of this interplay are not well understood. Here, we show that it readily leads to the emergence of two social morphs: a sessile, benevolent morph expressed by individuals who tend to increase the reproduction of others within their group relative to their own; and a dispersive, self-serving morph expressed by individuals who tend to increase their own reproduction. This social polymorphism arises due to a positive linkage between the loci responsible for dispersal and social behaviour, leading to benevolent individuals preferentially interacting with relatives and self-serving individuals with non-relatives. We find that this linkage is favoured under a large spectrum of conditions, suggesting that associations between dispersal and other social traits should be common in nature. In line with this prediction, dispersers across a wide range of organisms have been reported to differ in their social tendencies from non-dispersers.
Environmentally Mediated Social Dilemmas
Organisms modify their environments in ways that can be beneficial or detrimental not only to themselves but also to others sharing the same environment. Such niche-constructing or niche-destroying activities are often due to the production or consumption of environmental factors, such as resources, wastes, or toxins, which ultimately influence the ecology and evolution of social interactions. The authors present a new, four-way classification of social behaviors where individual behaviors are categorized into producing/consuming an environmental factor, as well as into helping/harming others.
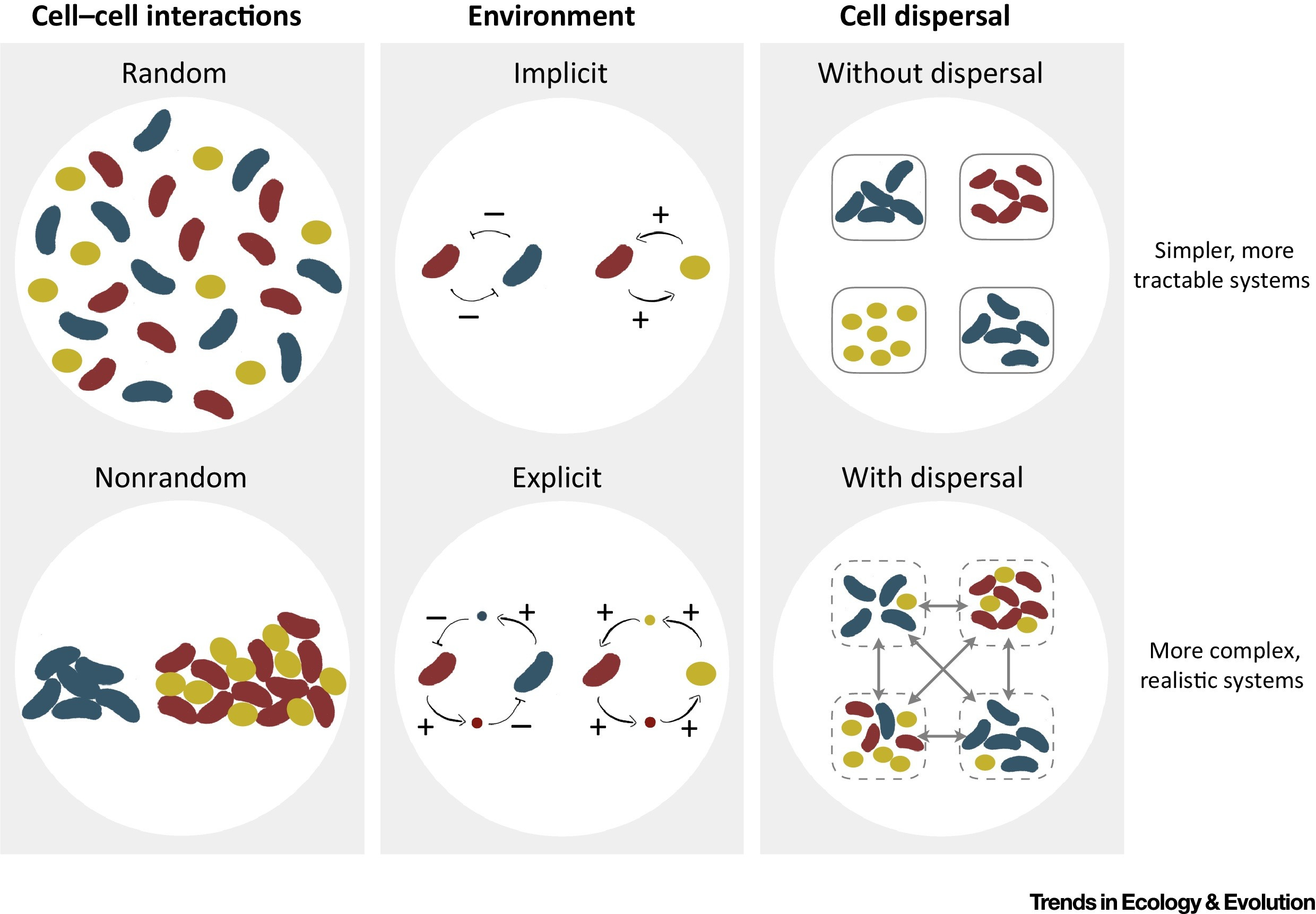
Although not immediately obvious, dispersal (the act of moving within a habitat or between habitats) is another mechanism by which organisms modify their environment. An explicit representation of such environmentally mediated interactions is key to capturing realistic system complexity and can reveal some unexpected outcomes in social dilemmas.
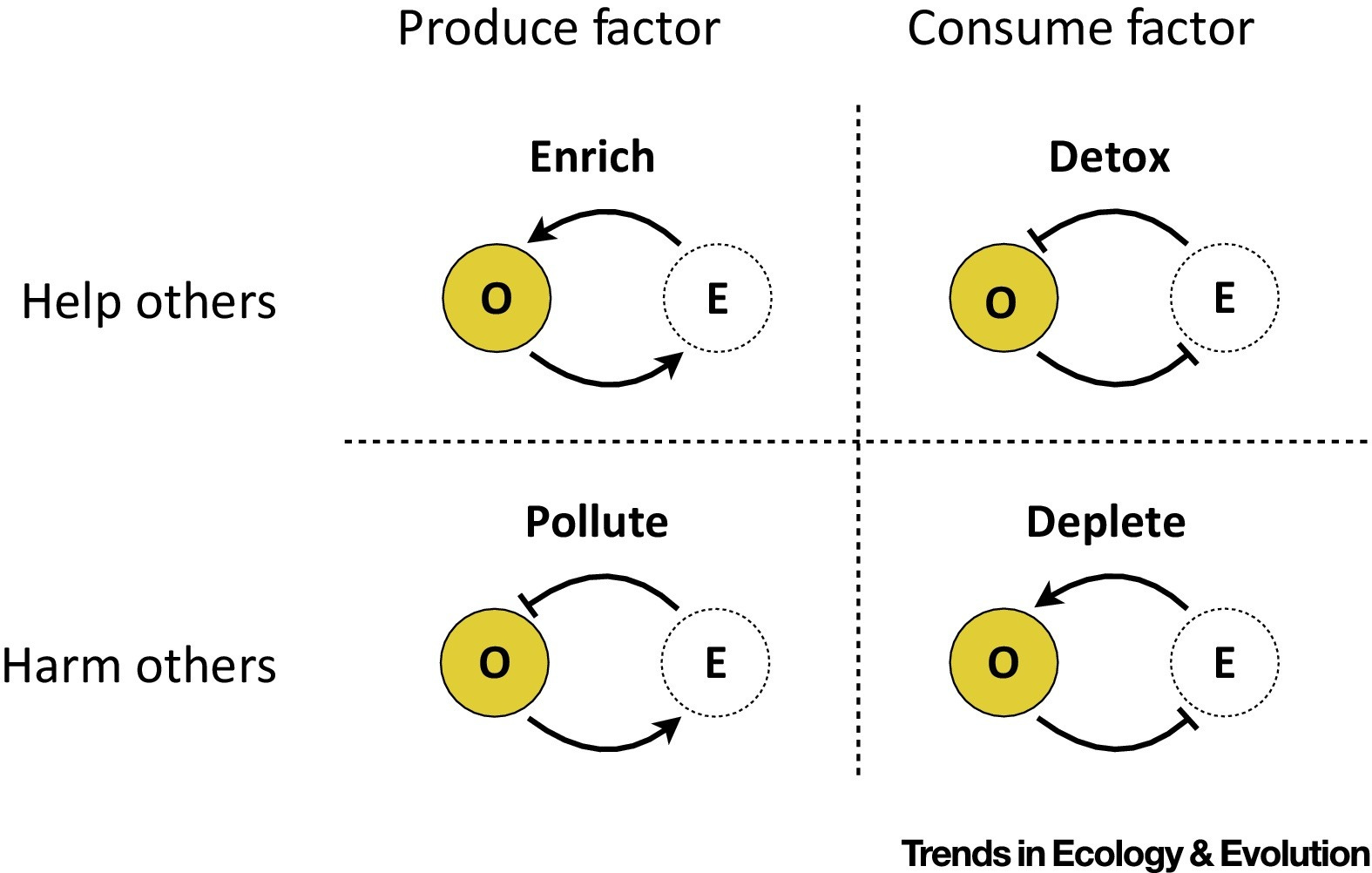
By consuming and producing environmental resources, organisms inevitably change their habitats. The consequences of such environmental modifications can be detrimental or beneficial not only to the focal organism but also to other organisms sharing the same environment. Social evolution theory has been very influential in studying how social interactions mediated by public ‘goods’ or ‘bads’ evolve by emphasizing the role of spatial structure. The environmental dimensions driving these interactions, however, are typically abstracted away. We propose here a new, environment-mediated taxonomy of social behaviors where organisms are categorized by their production or consumption of environmental factors that can help or harm others in the environment. We discuss microbial examples of our classification and highlight the importance of environmental intermediates more generally.
Ecosystems
A vast 4,000-year-old spatial pattern of termite mounds
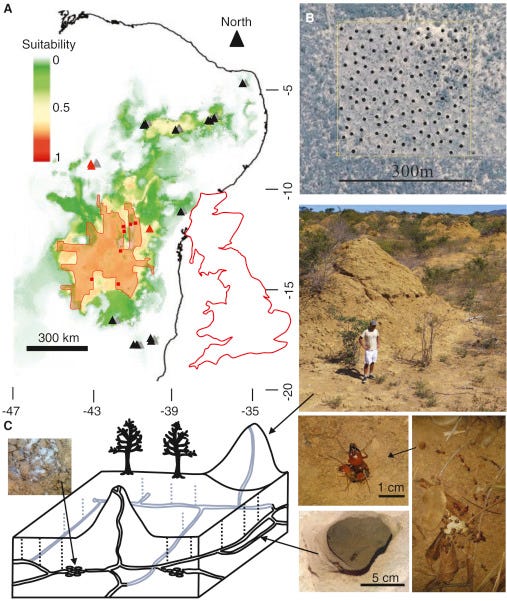
The origins of many large-scale ‘biogenic’ earthen structures are controversial, because often the species that built them have vanished. This is especially true when they form regular (over-dispersed), self-organized vegetation patterns [1]. Here, we describe a vast array of soil mounds constructed by termites (Syntermes dirus) that has persisted for up to 4000 years and covers an estimated 230,000 km2 of seasonally dry tropical forest in a relatively undisturbed and climatically stable region of Northeast Brazil. The mounds are not nests, but rather they are generated by the excavation of vast inter-connecting tunnel networks, resulting in approximately 10 km3 of soil being deposited in 200 million conical mounds that are 2.5 m tall and approximately 9 m in diameter. S. dirus termites are still present in the soil surrounding the mounds and we found that intra-specific aggression occurred at a scale much larger than an individual mound. We suggest that the complex network of tunnels built to access episodic leaf-fall has allowed for the optimization of waste soil removal, which over thousands of years has formed an over-dispersed spatial pattern of mounds.
Linking scaling laws across eukaryotes
Metabolic scaling theory has had a profound influence on ecology, but the core links between species characteristics have not been formally tested across the full domain to which the theory claims to apply. We compiled datasets spanning all eukaryotes for the foremost body mass scaling laws: metabolism, abundance, growth, and mortality. We show that metabolism and abundance scaling only follow the canonical ±3/4 slopes within some taxonomic groups, but across eukaryotes reveal reciprocal near ±1 slopes, broadly supporting the “energetic equivalence rule.” In contrast to metabolism, growth follows consistent ∼3/4 scaling within many groups and across all eukaryotes. Our findings are incompatible with a metabolic basis for growth scaling and instead point to growth dynamics as foundational to biological scaling.
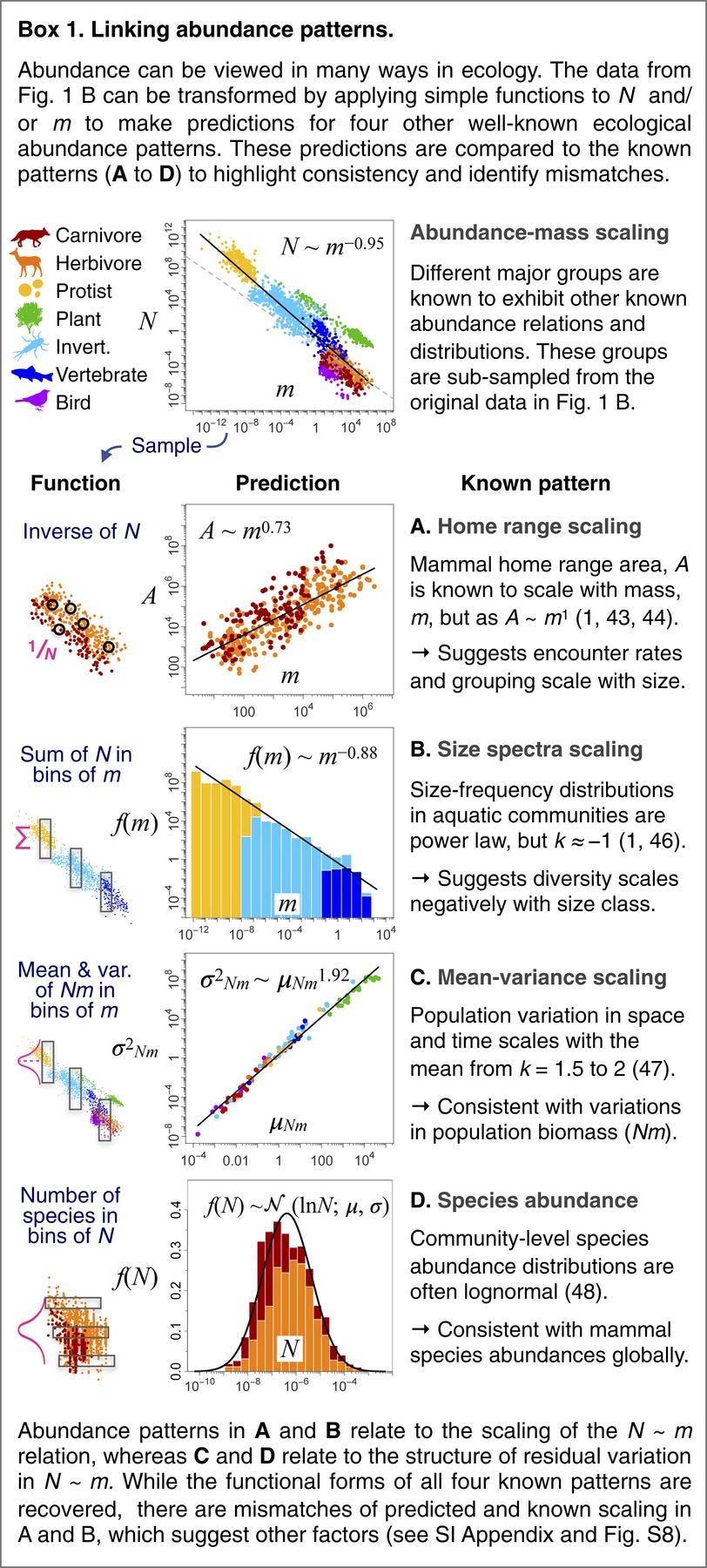
Scaling laws relating body mass to species characteristics are among the most universal quantitative patterns in biology. Within major taxonomic groups, the 4 key ecological variables of metabolism, abundance, growth, and mortality are often well described by power laws with exponents near 3/4 or related to that value, a commonality often attributed to biophysical constraints on metabolism. However, metabolic scaling theories remain widely debated, and the links among the 4 variables have never been formally tested across the full domain of eukaryote life, to which prevailing theory applies. Here we present datasets of unprecedented scope to examine these 4 scaling laws across all eukaryotes and link them to test whether their combinations support theoretical expectations. We find that metabolism and abundance scale with body size in a remarkably reciprocal fashion, with exponents near ±3/4 within groups, as expected from metabolic theory, but with exponents near ±1 across all groups. This reciprocal scaling supports “energetic equivalence” across eukaryotes, which hypothesizes that the partitioning of energy in space across species does not vary significantly with body size. In contrast, growth and mortality rates scale similarly both within and across groups, with exponents of ±1/4. These findings are inconsistent with a metabolic basis for growth and mortality scaling across eukaryotes. We propose that rather than limiting growth, metabolism adjusts to the needs of growth within major groups, and that growth dynamics may offer a viable theoretical basis to biological scaling.
Biological Systems
Approaching a state shift in Earth’s biosphere
Imagine tugging at a loose thread in a sweater: at first nothing much happens, but pull long enough and the whole fabric can unravel. Ecologists have long known that lakes, forests, or coral reefs can flip suddenly and irreversibly into entirely different states once pressures push them past a critical point. What is striking — and also deeply unsettling — is that the same may now be true for the planet as a whole.

Human activity has already transformed vast swaths of land, reshaped oceans with chemical runoff, altered the chemistry of the atmosphere and driven species toward extinction at unprecedented rates.
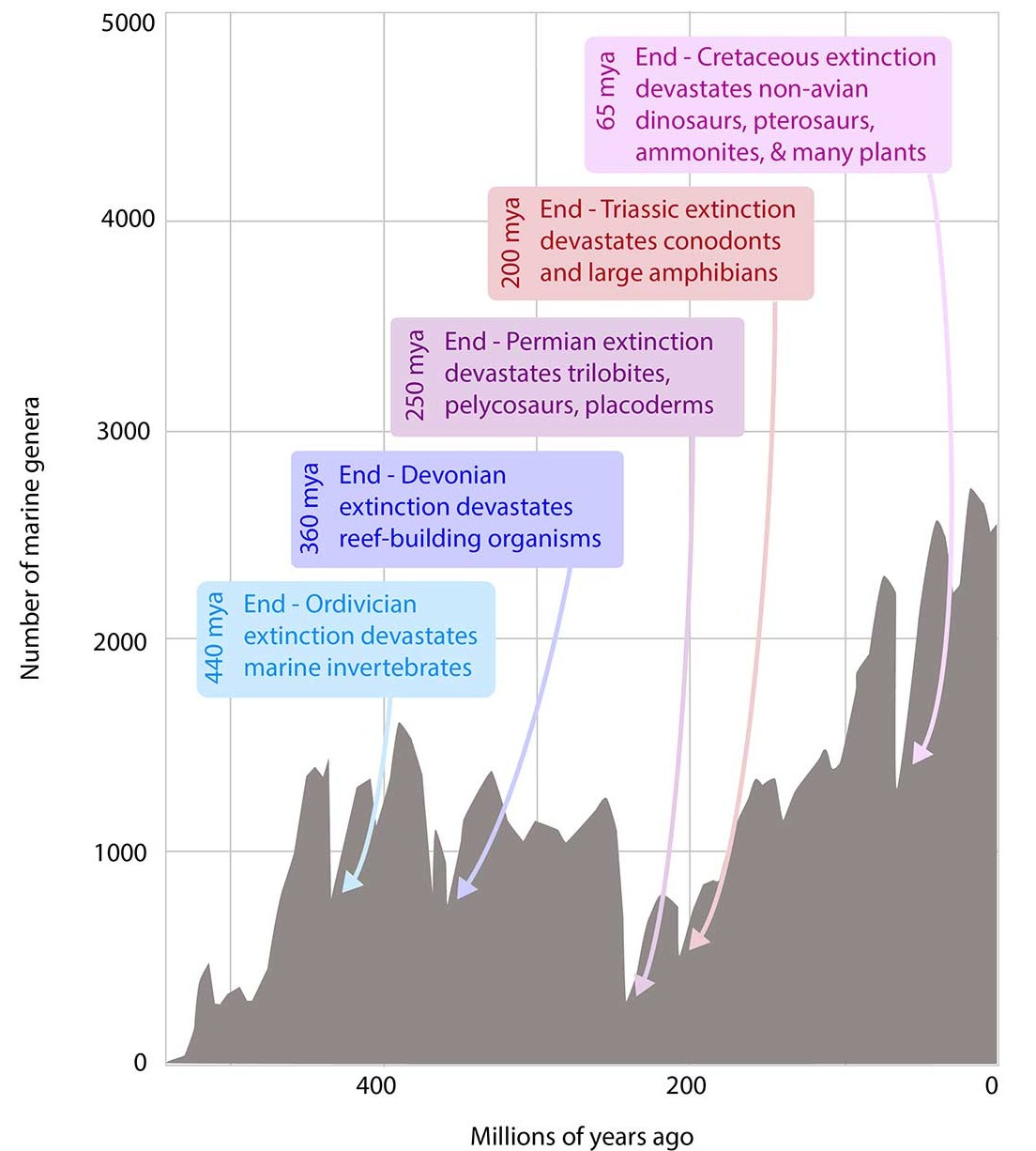
These local disruptions accumulate and interact, creating global-scale forces that can destabilize ecosystems even in remote places. The emerging picture is that Earth’s biosphere may be approaching a tipping point, where gradual pressures give way to abrupt planetary change. Recognizing this possibility is not just an academic exercise—it underscores the urgency of detecting early warning signs and addressing the root causes of environmental stress before we cross thresholds that cannot be reversed.
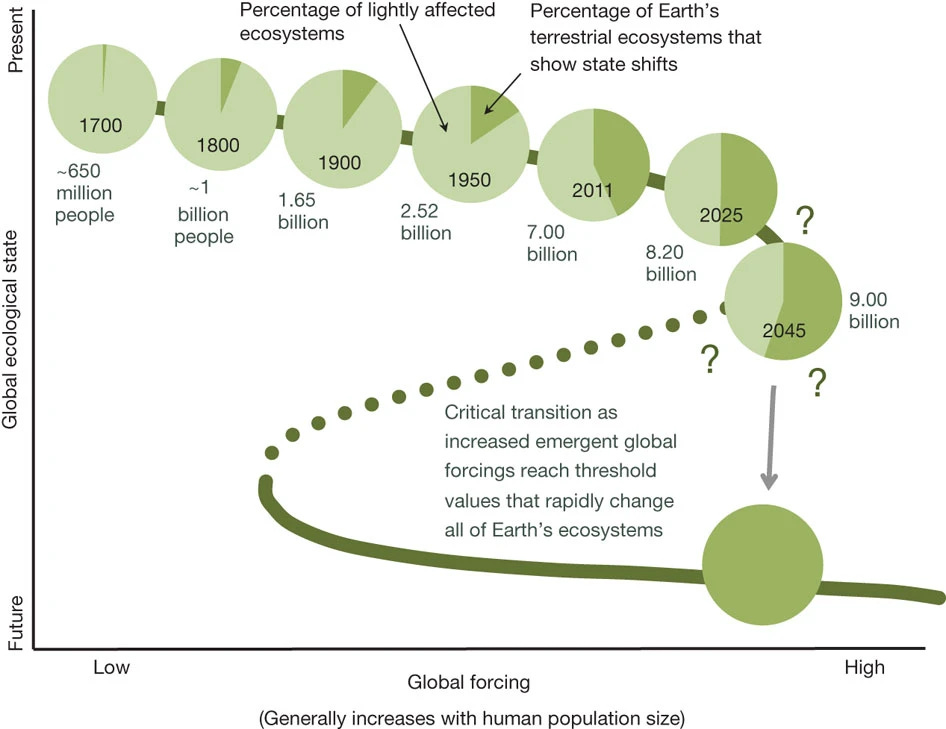
Localized ecological systems are known to shift abruptly and irreversibly from one state to another when they are forced across critical thresholds. Here we review evidence that the global ecosystem as a whole can react in the same way and is approaching a planetary-scale critical transition as a result of human influence. The plausibility of a planetary-scale ‘tipping point’ highlights the need to improve biological forecasting by detecting early warning signs of critical transitions on global as well as local scales, and by detecting feedbacks that promote such transitions. It is also necessary to address root causes of how humans are forcing biological changes.
→ Please, remind that if you find value in #ComplexityThoughts, you might consider helping it grow by subscribing, or by sharing it with friends, colleagues or on social media. See also this post to learn more about this space.