


From lone cells to organized organisms: multicellularity
From theory to practice and the importance of long-term experiments
If you find Complexity Thoughts interesting, follow us! Click on the Like button, leave a comment, repost on Substack or share this post. It is the only feedback I can have for this free service. The frequency and quality of this newsletter relies on social interactions. Thank you!
→ Don’t miss the podcast version of this post: click on “Spotify/Apple Podcast” above!
Is it possible to evolve multicellularity in the lab? What’s our understanding of the process and ability to reproduce it in silico and in vitro?
In one of their influential papers, John Maynard Smith and Eörs Szathmáry highlighted that life’s vast diversity can be traced (in principle) through a series of “major evolutionary transitions”.
Each transition — such as the move from free-floating genes to chromosomes, the jump from prokaryotes to eukaryotes, the rise of multicellular organisms and the emergence of sociality — represents a fundamental reorganization of how biological information is stored, shared and inherited, marking a turning point that has dramatically reshaped both the complexity of organisms and the rules of cooperation and conflict. By recognizing these specific leaps in life’s evolutionary history, they proposed a unifying lens for understanding how relatively simple beginnings could lead to the remarkable complexity we see in the biosphere today.
“There is no theoretical reason to expect evolutionary lineages to increase in complexity with time, and no empirical evidence that they do so. Nevertheless, eukaryotic cells are more complex than prokaryotic ones, animals and plants are more complex than protists, and so on. This increase in complexity may have been achieved as a result of a series of major evolutionary transitions. These involved changes in the way information is stored and transmitted.” — Eörs Szathmáry & John Maynard Smith (1995)
To overcome some of the drawbacks of this proposal, another paper from Szathmáry followed 20 years later. Nevertheless, some other issues still affect this re-organization of knowledge and you might be interested in reading a study attempting to enrich our understanding of macroevolution by including a metabolic perspective. I hope to find the required time to write more about this in another post.
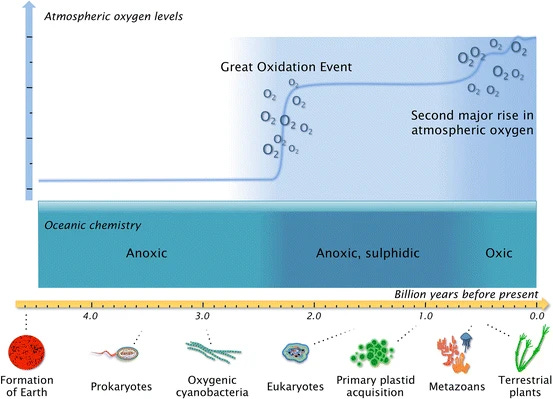
Regardless if we agree or not with the conceptual ground of the major evolutionary transitions, there is one specific phenomenon that we can surely describe as a critical change in the complexification of life. Multicellularity marks one of the deepest transitions in the history of life on Earth: it transforms independent, single-celled “individuals” into integrated, cooperative assemblages that share a common fate. This innovation unlocks new evolutionary possibilities, such as division of labor among cells and the emergence of complex structures like tissues, organs and nervous systems.
From an evolutionary standpoint, one of the key drivers behind multicellularity is the benefit of coordinated group living: single cells within a cluster can protect each other from predators, improve resource acquisition and buffer environmental changes, all advantages that could not easily achieved alone. However, multicellular organization also brings evolutionary trade-offs, including managing conflicts between cells and distributing nutrients efficiently. The balance of these forces has shaped an astonishing diversity of multicellular life forms, from simple algal aggregates to the intricate body plans of vertebrates.

If you are curious, check this great review by Knoll or this one by Niklas:
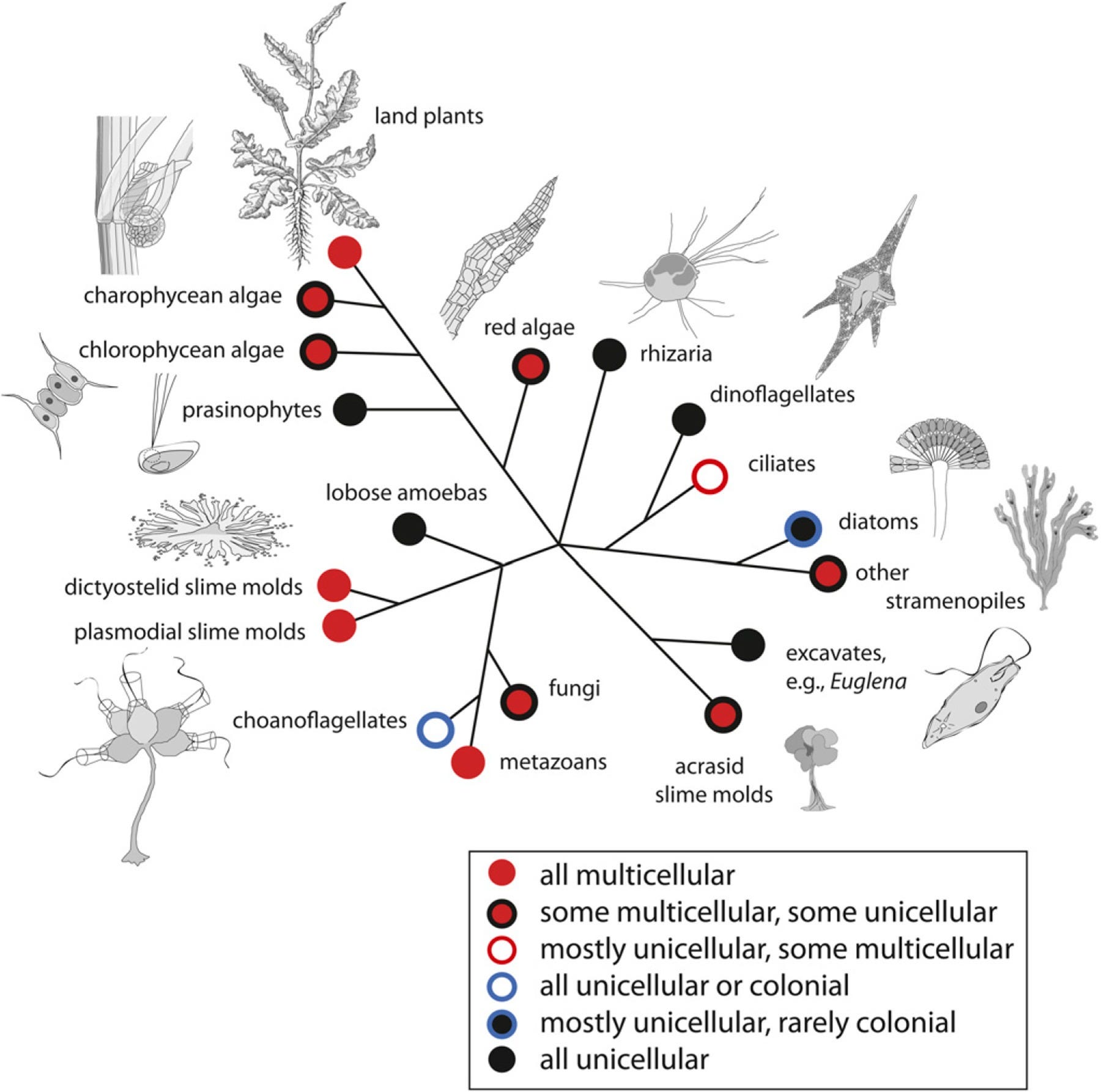
Multicellularity has evolved at least once in every major eukaryotic clade (in all ploidy levels) and numerous times among the prokaryotes. According to a standard multilevel selection (MLS) model, in each case, the evolution of multicellularity required the acquisition of cell–cell adhesion, communication, cooperation, and specialization attended by a compulsory alignment-of-fitness phase and an export-of-fitness phase to eliminate cell–cell conflict and to establish a reproductively integrated phenotype. These achievements are reviewed in terms of generalized evolutionary developmental motifs (or “modules”) whose overall logic constructs were mobilized and executed differently in bacteria, plants, fungi, and animals. When mapped onto a matrix of theoretically possible body plan morphologies (i.e., a morphospace), these motifs and the MLS model identify a “unicellular ⇒ colonial ⇒ multicellular” transformation series of body plans that mirrors trends observed in the majority of algae (i.e., a polyphyletic collection of photoautotrophic eukaryotes) and in the land plants, fungi, and animals. However, an alternative, more direct route to multicellularity theoretically exists, which may account for some aspects of fungal and algal evolution, i.e., a “siphonous ⇒ multicellular” transformation series. This review of multicellularity attempts to show that natural selection typically acts on functional traits rather than on the mechanisms that generate them (“Many roads lead to Rome.”) and that genome sequence homologies do not invariably translate into morphological homologies (“Rome isn't what it used to be.”).
Reproducing multicellularity in the lab
While the concept of multicellularity can seem abstract at first sight, modern experimental evolution allows scientists to watch it unfold in nearly “real time”. By creating environments that favor group-level survival, researchers can push single-celled organisms to evolve cooperative, multicellular traits. For instance, controlled settings might reward larger cluster size (i.e., cells that stick together are more likely to survive).
Over many generations, traits that support cohesion, communication and division of labor emerge naturally, as shown in a paper by Will Ratcliff and colleagues more than one decade ago. These experiments are amazing because they can be thought as small-scale “time machines” that reproduce millions of years of evolution into a matter of weeks or months. However, at variance with natural evolution out of the lab, such experiments provide us with precise control of the settings and monitoring, letting researchers manipulate factors like nutrient availability or predation pressure to see how the balance of cooperation and conflict evolves under different conditions.
Will, professor at Georgia Tech and Director of QBioS, is a leading figure in this experimental domain, and his seminal work with yeast (specifically, the “snowflake yeast”) has provided a living model of how single-celled organisms can transition to a simple form of multicellularity under laboratory selection.

Personally, I have discovered Will’s work when I was investigating a network model (still under development) of multicellularity. At that time I asked him to re-use the above figure and, at the same time, if he could guide me towards existing network-based approaches to multicellularity. The funny thing was that he pointed to one of my own papers, a recent work I did with Ricard Solè and several other stellar collaborators from Santa Fe Institute and other institutions (you can read a dedicated post following the link below). I got that I was on the right track.

In Will’s experiments, yeast populations were repeatedly subjected to conditions favoring faster-settling clusters: accordingly, within a few dozen generations, many yeast lineages had evolved the tendency to remain attached to their offspring, forming branching, tree-like clusters (like in the above figure). This cluster-level organization paved the way for characteristics reminiscent of multicellular life: coordinated growth, “cheater” suppression mechanisms and rudimentary forms of cell differentiation. I also find extremely cool that Will’s lab produces kits allowing you to perform your own multicellular yeast evolution experiment or run a predation experiment!
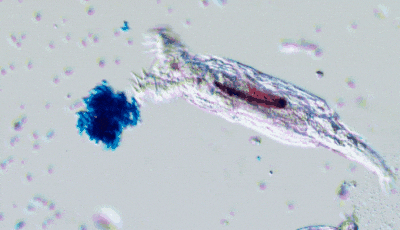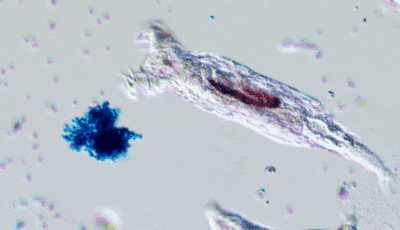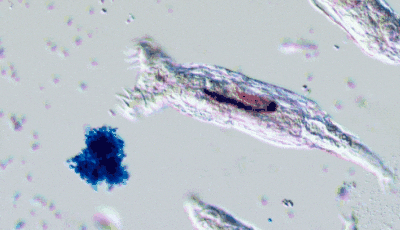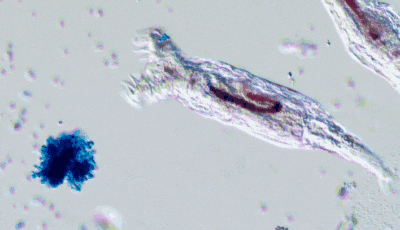
In a recent review paper with James Stroud, they highlight the importance of long-term experiments to observe evolutionary phenomena that would not be observable otherwise:

I have invited Will to #ComplexityThoughts to answer some pressing questions and better understand his perspective on this matter.
Can you tell us more about how did you come with the idea of playing time machines in the lab to force specific evolutionary pathways?
The great strength of evolutionary biology is the comparative record—we know multicellularity has evolved repeatedly across the tree of life, yet in every case those transitions are ancient. This creates a fundamental challenge: connecting microevolutionary dynamics to macroevolutionary patterns is difficult because we can't examine how multicellularity originated in real-time. Laboratory evolution provides insight into this dynamic process.
What fascinates me is how cells transition from being primary units of selection—gaining adaptations as single-celled organisms—to surrendering their individuality and becoming parts of a new higher-level organism. This step is especially hard to detect in lineages where it occurred billions of generations ago.
I should clarify that I don't view our approach as forcing specific evolutionary pathways. The Multicellularity Long Term Evolution Experiment (MuLTEE) is designed to be open-ended regarding how yeast solve major challenges in multicellular evolution. We're not dictating specific pathways but rather presenting evolutionary challenges and allowing them to discover solutions through multicellular adaptation. This creates a unique window into evolutionary processes that we couldn't otherwise observe.
How far do you think experimental evolution can push micro-organism clusters toward more sophisticated multicellularity?
That's a fascinating question without a definitive answer—we simply won't know until we try. There are no comparable laboratory evolution experiments to serve as benchmarks because, to my knowledge, no one has attempted to evolve entirely new kinds of organisms over truly long-term evolutionary timescales.
If we take prior evolution as our guide, there are essentially no inherent constraints. The evolution of multicellularity is an open-ended process in which, theoretically, we could push a laboratory system as far as any natural system has gone. While I don't expect we'll evolve plants, animals, or mushroom-forming fungi—sophisticated organisms that evolved over vast timescales—I don't see any intrinsic limitations to the process.
The main constraints will be practical: time and our ability to identify and leverage the right evolutionary mechanisms. The MuLTEE provides an unprecedented opportunity to observe thousands of generations of adaptation, but even this represents just a moment in evolutionary time compared to the hundreds of millions of years that shaped complex multicellular life on Earth.
Which do you find more critical in maintaining stable multicellular structures: altruistic cooperation or effective conflict management?
Neither. I think the social evolution field has convinced us that cooperation and altruism are the greatest challenges for multicellularity to overcome, but I consider this a relatively trivial problem for most multicellular systems.
It's easy to avoid conflict by forming groups with clonemates, which is easily accomplished via genetic bottlenecks in the life cycle. Conflict in these lineages only becomes problematic in very large, long-lived organisms where mutation generates enough diversity during an organism’s life time for within-group selection to arise. While that's relevant for organisms like us, cancer-like dynamics are less important for other multicellular lineages. Even when cancer-like dynamics emerge in plants or fungi, they're typically not catastrophic since these modular systems have cells connected by permanent bonds that prevent cancerous lineages from spreading throughout the organism. Also, a tree losing a branch is usually much less fatal than an animal losing a heart or brain.
I believe the major challenges in evolving complex multicellularity aren't about cooperation and conflict but rather mutual information and coordination—the fine-tuning of complex morphogenesis. This perspective shifts our focus from how complexity is eroded through social conflicts to the opposite side of the coin: how complexity arises through the evolutionary construction of integrated developmental systems that enable sophisticated collective-level adaptations.
Were you surprised by how quickly your yeast populations evolved group-level traits in the lab?
Absolutely, though we weren't met with instant success. We attempted several early versions of the MuLTEE that failed. Our populations hit adaptive plateaus with no further multicellular evolution, leaving me uncertain about continuing the experiment.
The breakthrough came when Ozan Bozdag, then a postdoc and now a research scientist in my lab, tried different metabolic treatments that modified oxygen availability. We discovered that oxygen acts as a constraint on size when it's a valuable resource. However, when you overcome this limitation—either by providing extra oxygen or removing the need for oxygen in metabolism—you eliminate a critical constraint on growth, enabling open-ended multicellular adaptation.
This opened fascinating new evolutionary possibilities. As clusters grow larger, they encounter different problems with resource diffusion, creating opportunities to break trade-offs through evolving cell differentiation, circulation systems, and novel resource provisioning behaviors. In this sense, I've been genuinely surprised by how quickly our yeast are evolving novel multicellular innovations once key constraints are removed.
Could the same experimental principles used with yeast work for other, more complex organisms? What are the limits that we can reach with this technology? And how might that expand our understanding of multicellularity?
I don't see why not, though we won't know until we try. I would love to see others apply long-term experimental evolution to explore new kinds of organisms. For example, there's no fundamental reason why one couldn't take a solitary bee species and evolve it toward becoming a eusocial superorganism through appropriate selection regimes. Or explore the boundaries of super-organismality among plants.
Importantly, this approach isn't limited to going ‘up’ the ladder of biological complexity. There are tremendous opportunities to investigate earlier transitions in individuality as well. I'd be fascinated to see this methodology applied to origin-of-life questions, the emergence of cellular life, or the origin of eukaryotes through symbiosis between previously free-living prokaryotic lineages.
The MuLTEE demonstrates that experimental evolution can reveal fundamental principles governing major evolutionary transitions. By observing these processes unfold in real-time, we gain surprising and unique insights that complement what we know from the comparative record and evolutionary theory. The limits of this technology remain unknown, but we’re just scratching the surface of what it can teach us about evolutionary innovation and the emergence of biological complexity.
And we will wait for Will’s lab new results about this!
Meanwhile, in silico modeling of the emergence and evolution of multicellularity is a highly interdisciplinary challenge that combines evolutionary theory, developmental biology and computational science. Honestly, I think that there is room for statistical physics and dynamical systems theory to join. Check below some of the ongoing efforts in this direction, bridging current questions with concepts proposed in 1940s and later made popular in the 1970s-90s.
Thank you for this wonderful blog tackling these deep questions. I love it.......But, but.... What if yeasts had evolved millions/billions of years before "our" solar system had evolved? And yeasts are widely distributed throughout the Universe.... or at least "our" galaxy. Including in meteorites or even other planets like Mars for example. And earth was seeded by multicellular, sexually reproducing organisms, perhaps on multiple occasions, by hitching a ride on solid space debris? None of this "evolution" of either single cellular or multicellular life forms actually occurred on the planet "Earth"?