
What network structure alone cannot tell us about complex systems
Structure shows where interactions can happen, while functionality depends on how perturbations actually move
If you find value in #ComplexityThoughts, consider helping it grow by subscribing and sharing it with friends, colleagues or on social media. Your support makes a real difference.
→ Don’t miss the podcast version of this post: click on “Spotify/Apple Podcast” above!
A network map can show how a system is wired, but — at least, in general — it does not tell us what the system can still do when disturbed. To understand functionality, we have to follow perturbations, flows, time scales and the diversity of possible responses.
This post is adapted from a seminar I recently gave in May for the Making Connections series hosted by the University of Cambridge and King’s College London. Since the seminar lasts about one hour, and we live in the 8-seconds era (I will write more about this in another post), I thought that it could be helpful for you to have a 8-minutes summary.
The aim of the seminar was to explain, in a not-too-technical way, a framework my group has been developing to study network functionality in complex adaptive systems. I realize that it is a long story, after all, since my first papers about this matter have been published in 2016 and 2017. The question is simple to state:
Where do the “superpowers” of complex systems come from?
To this aim, let us consider some emblematic examples:

a tardigrade, which is a microorganism able to survive extreme temperatures, radiation, pressure and dehydration.
an axolotl, which is capable of regeneration.
the fruit fly connectome, where information-processing architecture supports behavior.
a slime mold, which has no nervous system but can solve spatial exploration problems by building efficient networks.
fire ants, which during floods form a living floating platform without central coordination.
This is not limited to biological systems, though. Take the Internet, a system that continuously changes state: route announcements, flaps, validation, outages, path hunting, mitigation. The dynamics running through the network is the most relevant information, here. But if global connectivity coupled to decentralized routing and continual perturbations predicts fragility, why does the Internet remain functional most of the time?
These examples differ in substrate, scale and biological organization, with some capacities being genetically encoded, whereas others emerging from collective interaction (eg. in ants). Yet they raise the same scientific problem: how can a system acquire a repertoire of behaviors that allows it to survive, adapt, regenerate, process information or coordinate under perturbation? This is also the core question of my seminar and, more generally, of several of the research activities in my Lab.
In fact, network science gives us part of the answer, but only part. A network can describe how genes regulate genes, how metabolites interact, how neurons connect, how individuals coordinate, or how infrastructures are linked. In a system such as C. elegans (a nematode usually adopted as model organism), one can represent genetic regulation, metabolism and the connectome as different layers of organization. In living systems more generally, one can move across scales: DNA, proteins, pathways, cells, organisms, populations.
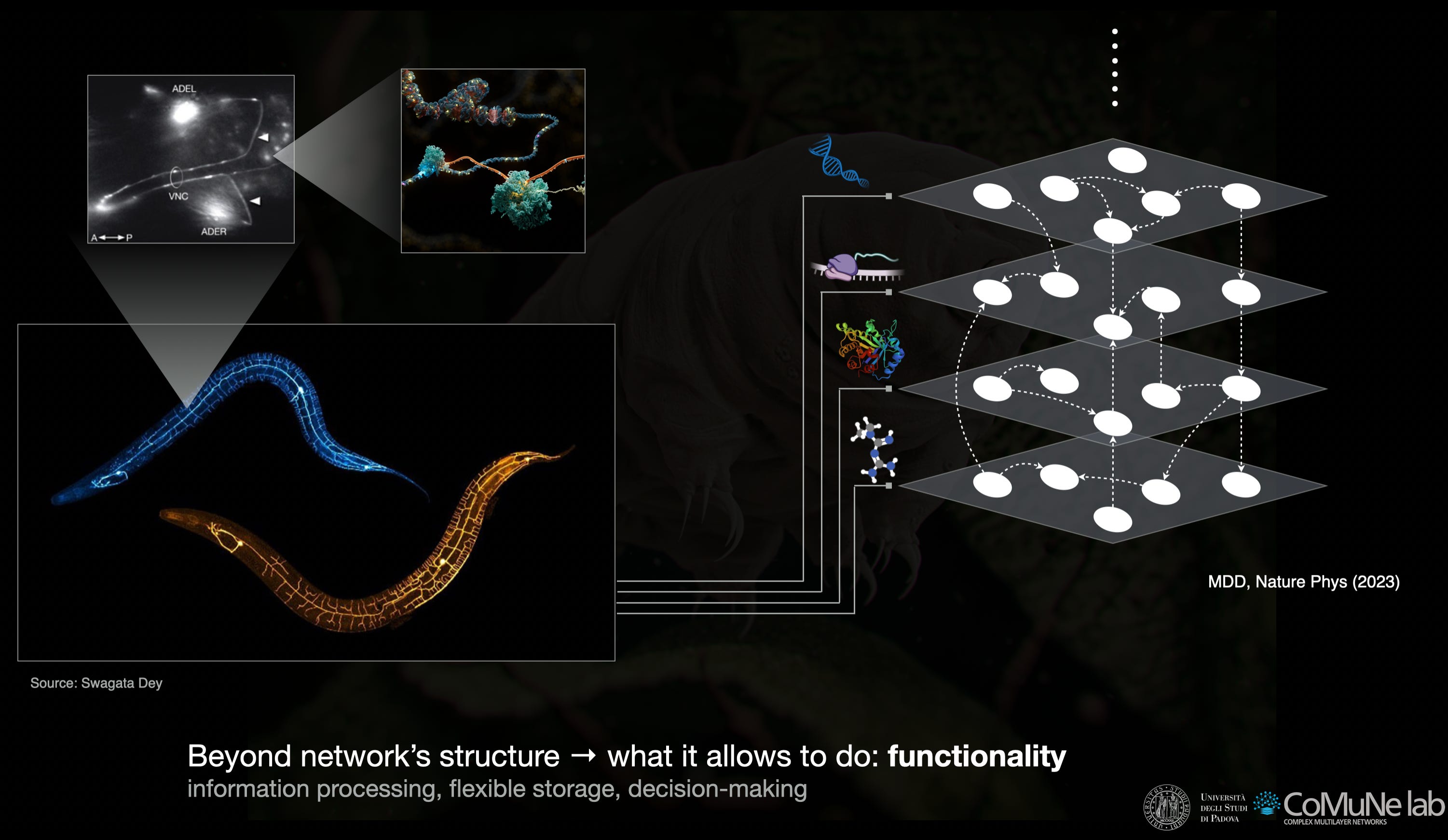
This structural information is essential, but it is not the same as functionality. For instance, let us consider a drug: it acts at the molecular level, but its relevant effects may appear at higher scales. Similarly, a perturbation introduced in one part of the system can propagate elsewhere, sometimes (but fortunately not always, especially in biology) across levels of organization. While structure constrains these processes, it does not by itself determine what the system can do.
A useful way to see the distinction is an infrastructure example. In the seminar I showed data from the New Orleans outage in 2021. The city remained there: its streets, neighborhoods and spatial layout did not disappear. Yet the functionality supported by that structure was impaired. The map of the system survived, whereas some of the flows that made the system work did not.
This is the central point: functionality is not reducible to structure. But what is functionality? Let me define it more operationally.
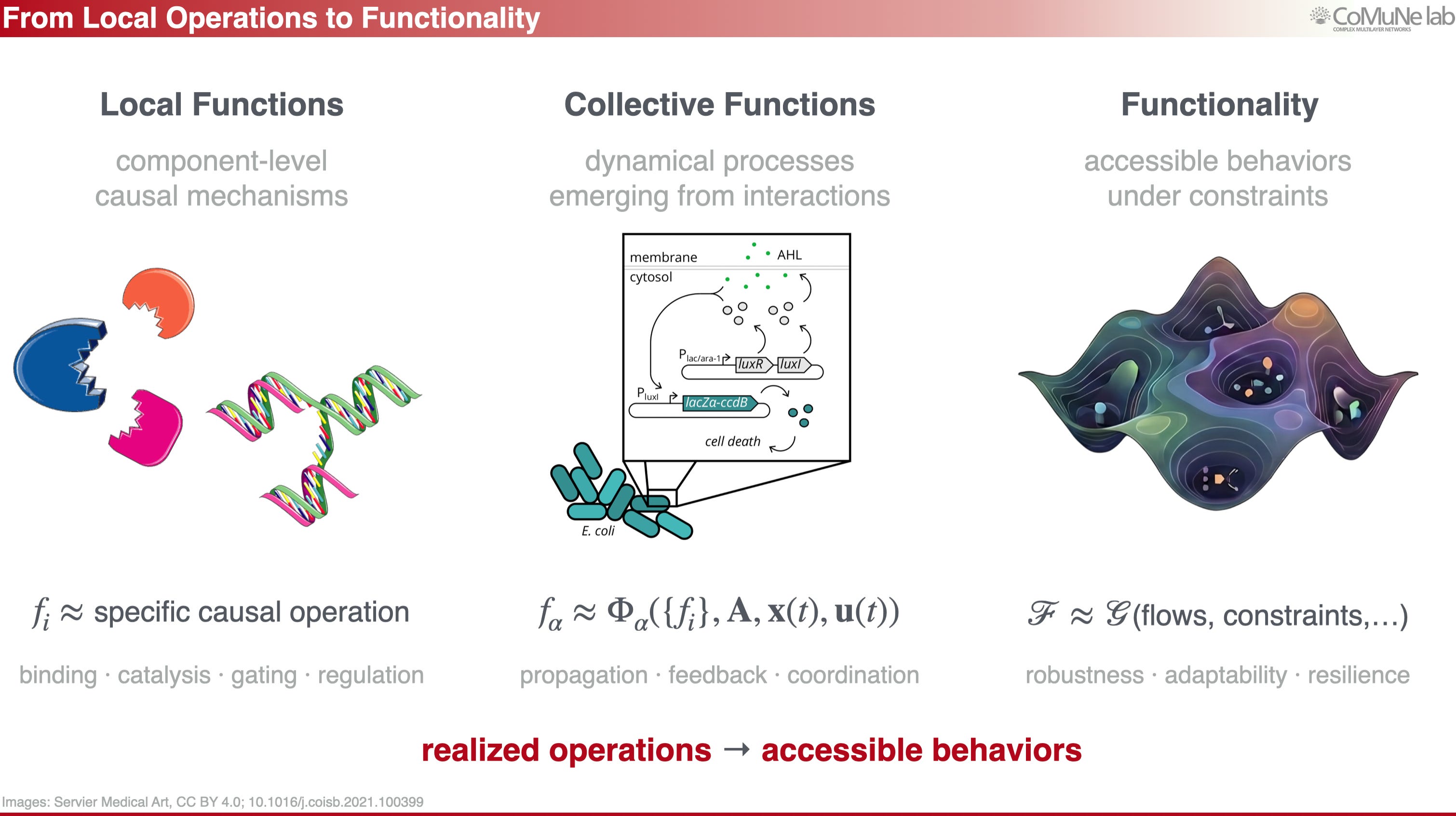
A function is a realized operation, such as binding, catalysis, gating, signal propagation, or coordination. Functionality is broader: it is the set of accessible behaviors a system can express under given constraints. Robustness, adaptability and resilience are widely known examples of functionality, because they describe what a system can still do when conditions change.
The methodological step
Scientists do not observe reality directly: we observe filtered traces of it, at best. Our measurements select variables, instruments impose limits and models translate observations into hypotheses about mechanisms.
In the seminar I used flocking as an example: birds form decentralized collective motion, but a simulation can reproduce essential features of that phenomenon without using biological birds as the substrate. What matters here is not only what the system is made of, but the generative rules that can reproduce the observed behavior.
This is why complexity science needs generative models: they provide a compressed account of cause and effect, when cause and effect can be meaningfully specified. To build such models in a principled way, in physics we often relies on variational principles:
define a quantity → impose constraints → find the state/trajectory that extremizes it
Stationary action provides one classical example of this procedure, maximum caliber extends this logic to ensembles of stochastic trajectories, while maximum entropy applies when one has constraints on steady states rather than full trajectory information.
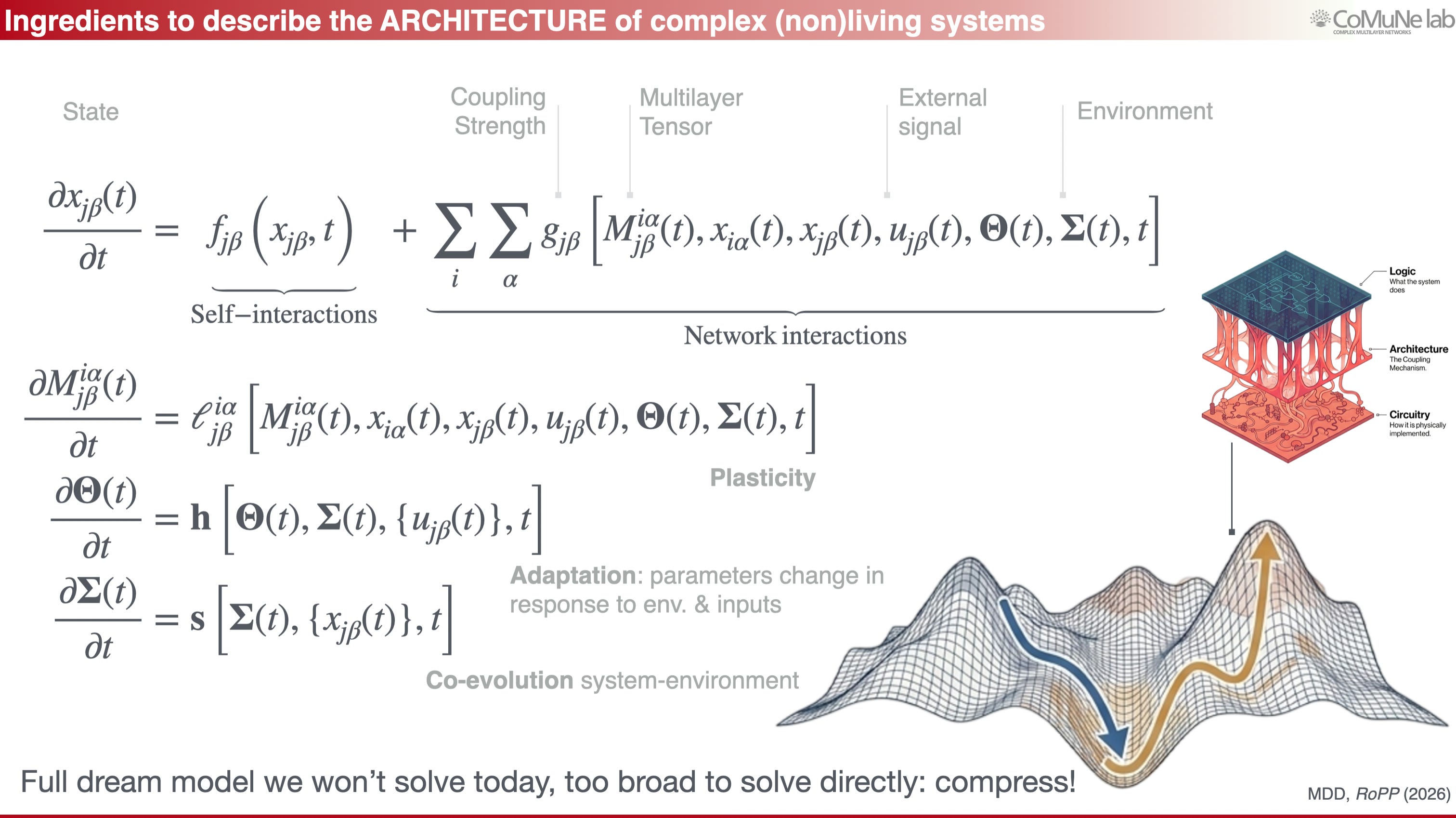
For network functionality, the ingredients are analogous, but there are important differences. Roughly speaking, we need:
a statistical description;
constraints linking microscopic behavior to macroscopic observables;
an entropy measure.
However, the statistical object in our case cannot be an usual ensemble of networks: it is an ensemble of perturbation propagation events over a network. This shift is the core of the framework.
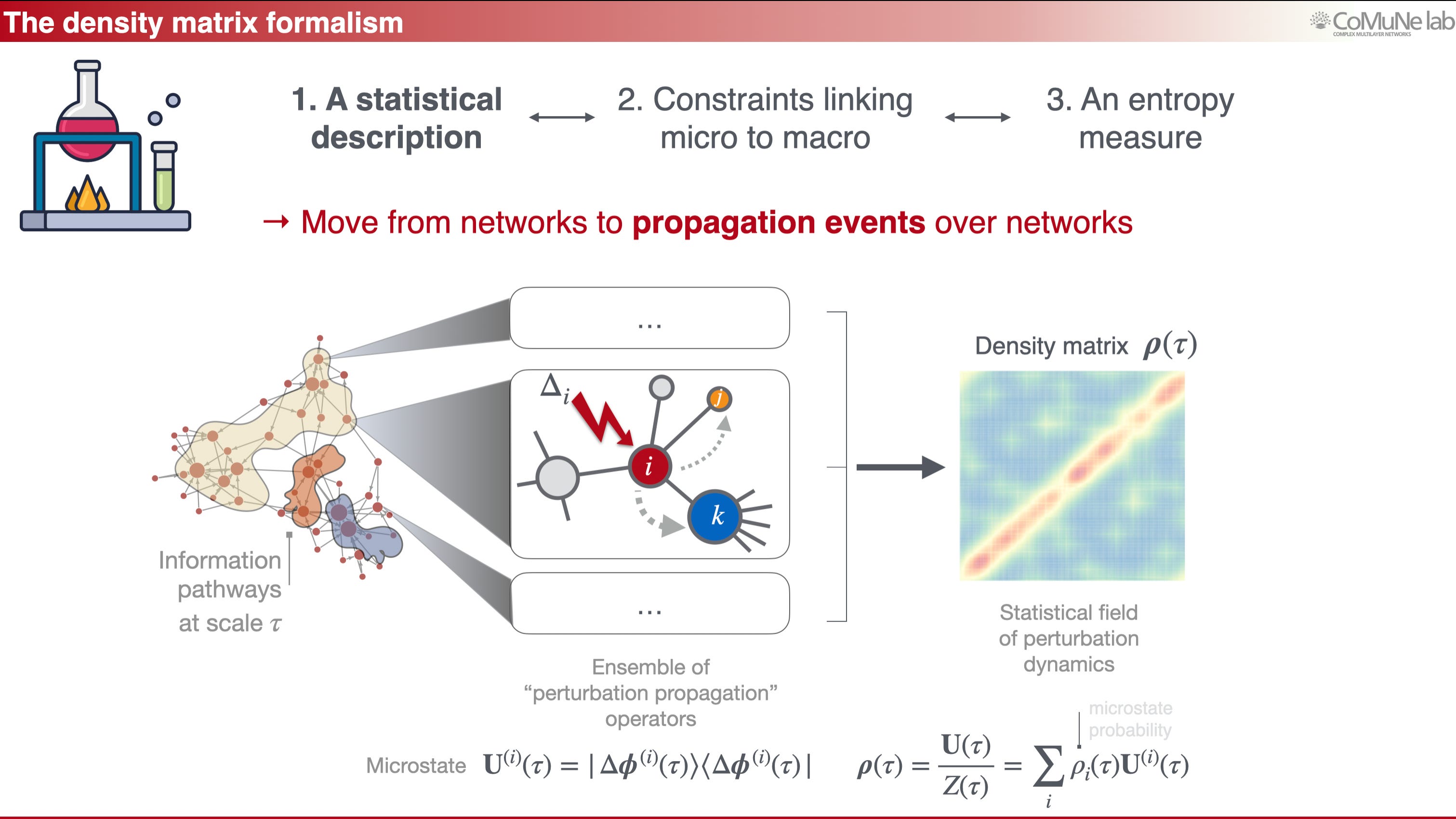
Imagine perturbing one node of a system at a given time scale. The perturbation may spread locally, reach a mesoscale module, or eventually influence global organization. If we consider all possible perturbations and the pathways they activate, we obtain a statistical description of information propagation. This can be encoded in an operator, a density matrix, which captures the interplay between structure and dynamics.
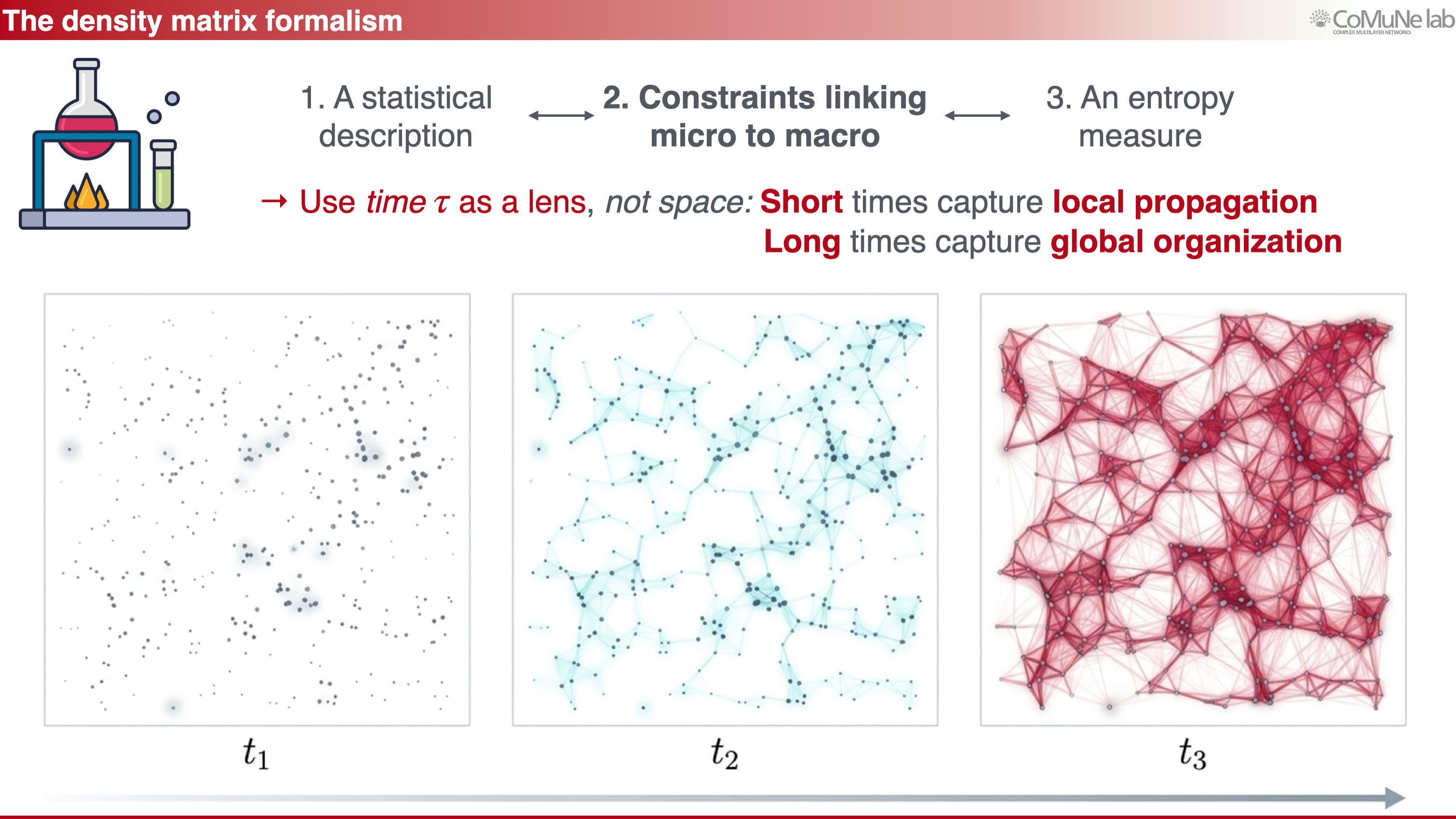
Time then becomes a lens: at short time scales, one sees local response; at intermediate time scales, mesoscale organization appears; at long time scales, components and global integration become visible.
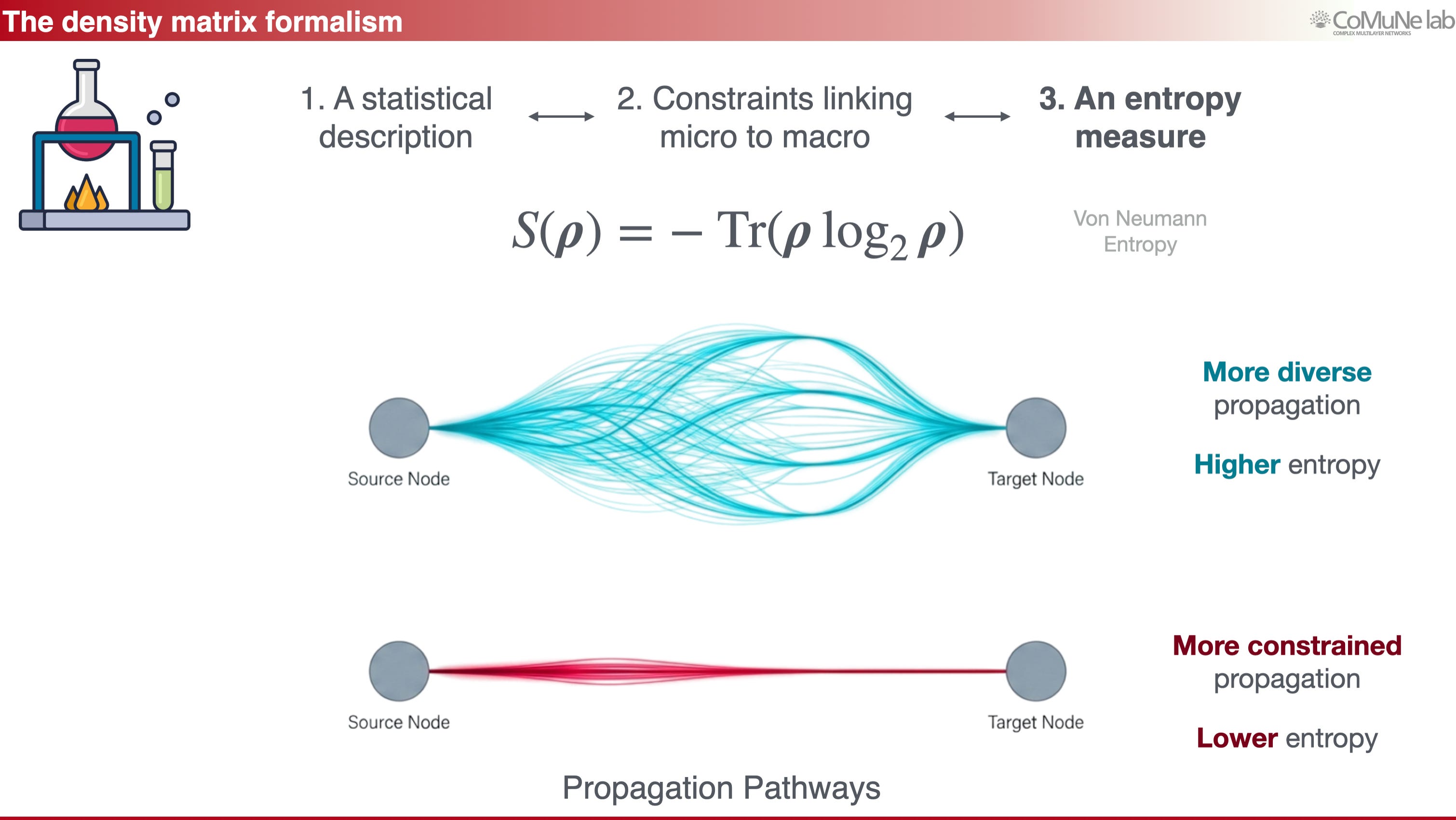
From a mathematical point of view, our language resembles quantum mechanics, even though the interpretation is very different. The density matrix is used because it provides the right mathematical object for describing propagation ensembles. Its von Neumann entropy measures the diversity of responses to perturbation: high entropy means that perturbations can explore many pathways, whereas low entropy means that propagation is constrained.
Functionality is therefore not a single number attached to a network; it cannot be: it depends on perturbation, dynamics and observation scale.
Applications
First, functionality can break even when structure does not collapse. Classical network robustness often studies attacks by asking whether the largest connected component survives. This is useful, but it is based on the assumption that structural degradation as a proxy for functional degradation. In fact, it is easy to show why this proxy can fail: if two subsystems need to exchange flow at a relevant time scale, a local attack may leave the network mostly connected while delaying or suppressing that exchange over short-to-mid time scales. In a connectome, for example, a local perturbation may preserve much of the wiring while degrading information transmission over biologically meaningful times. This motivates functional importance measures: for instance, remove or perturb a node, recompute propagation entropy, and rank nodes by their effect on information flow.
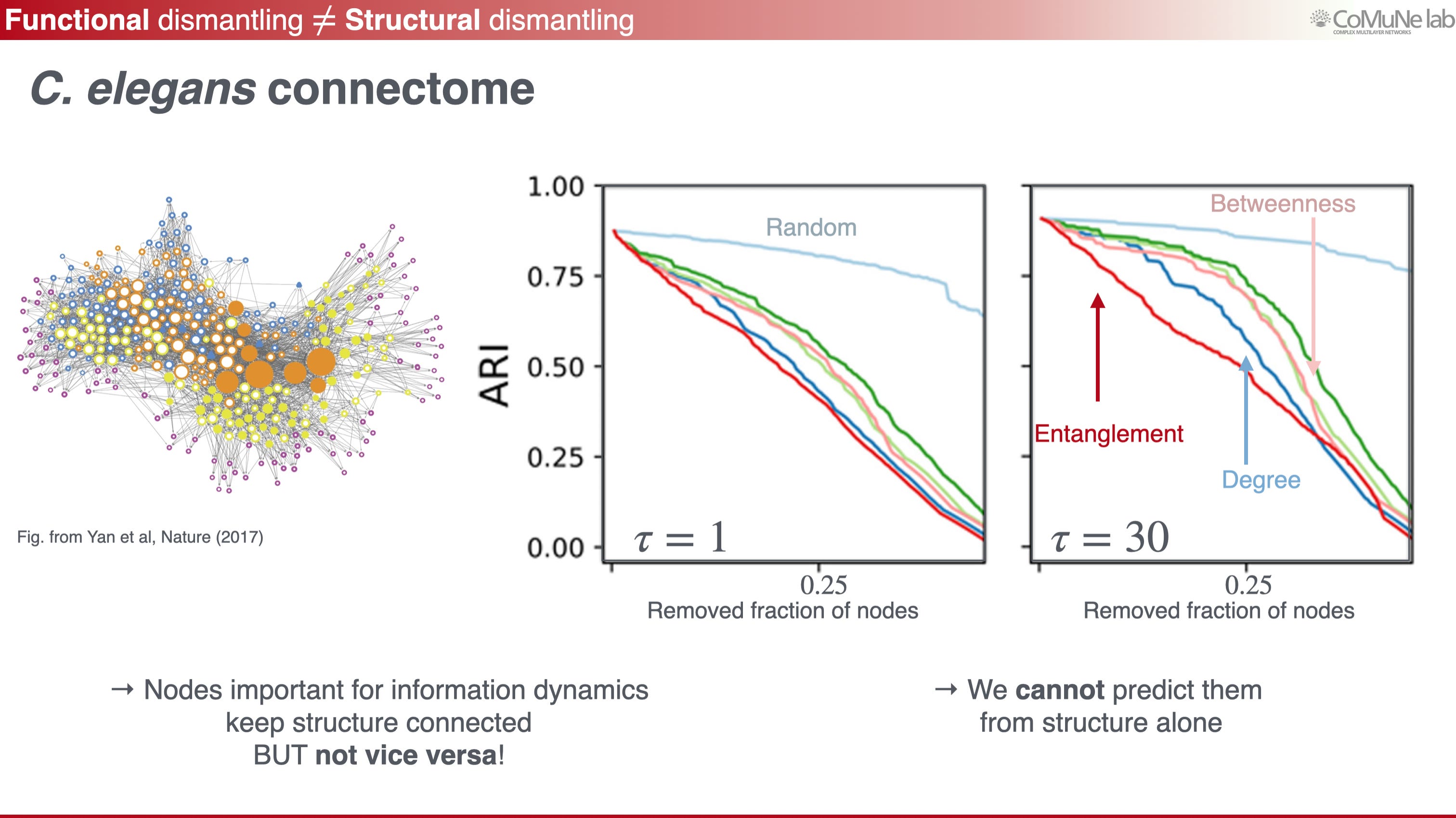
In applications to C. elegans and human connectomes, nodes that are important for functional propagation are not necessarily the same as nodes that look important from structure alone (see below, and assume that ARI is a measure of how good the network is at functional):
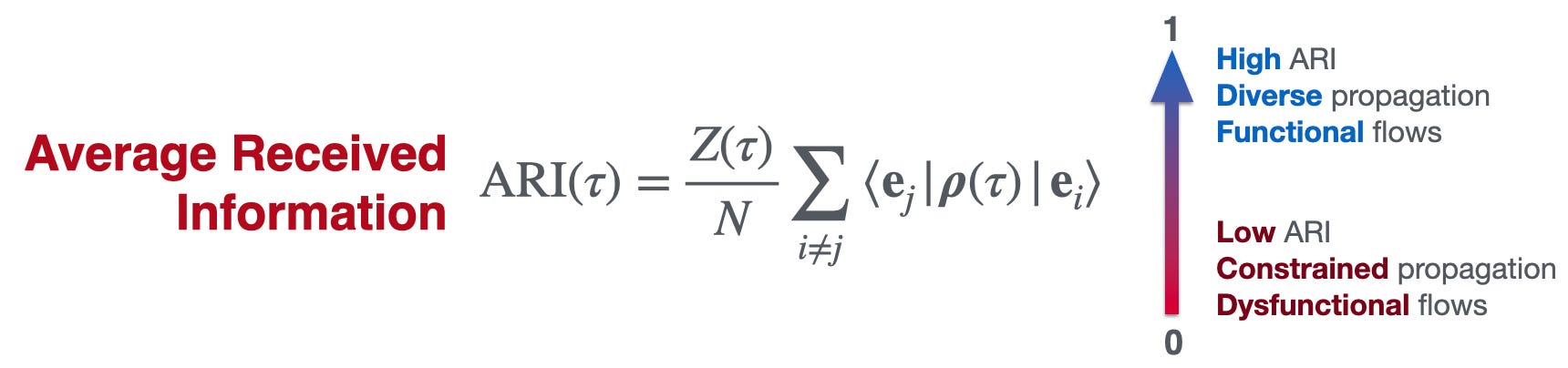
Second, functionality involves a thermodynamic trade-off:
A nearly disconnected network has too few links to support global propagation;
A fully overconnected network spreads perturbations too broadly, so responses lose diversity.
Between these regimes lies a functional balance: signals can travel efficiently, but the system does not respond everywhere in the same way.
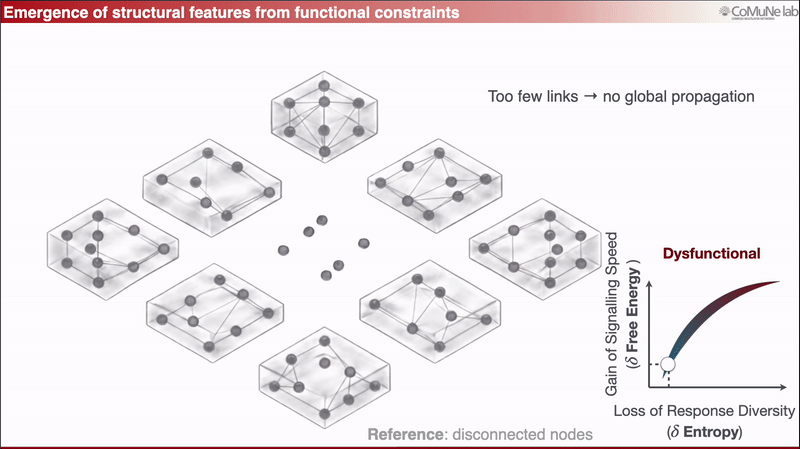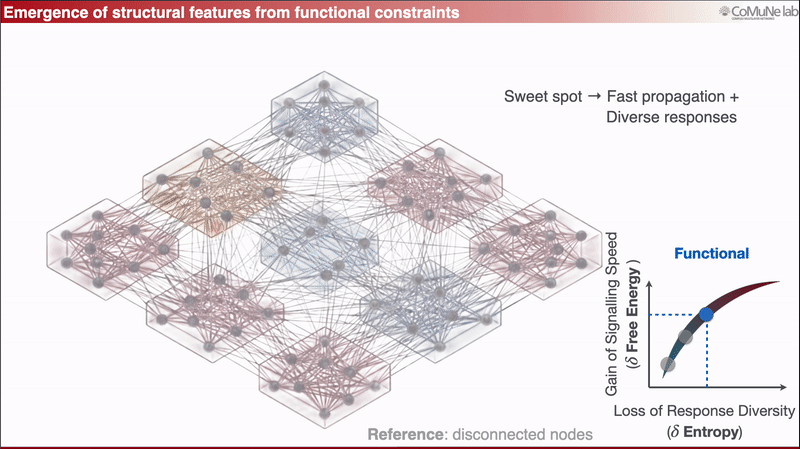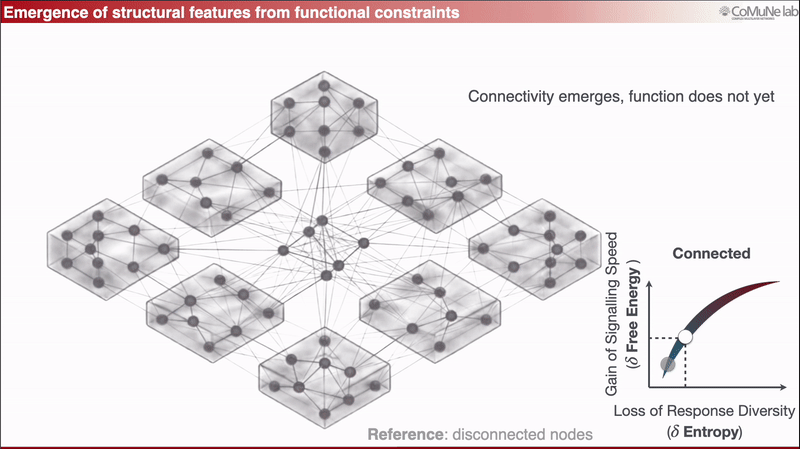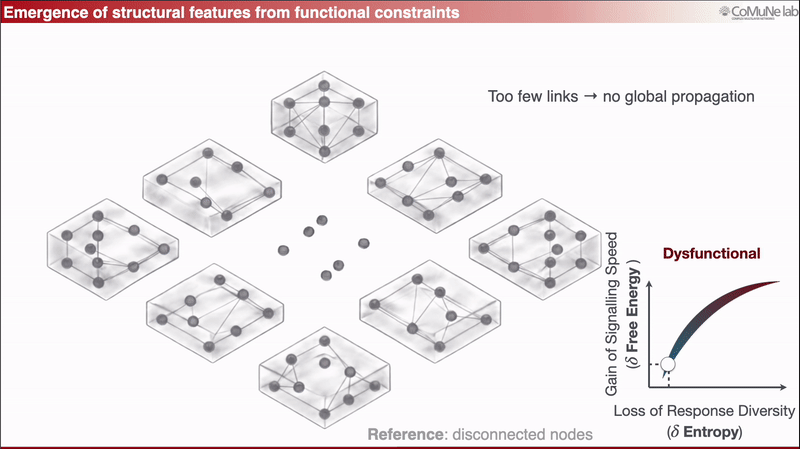
This can be expressed through an efficiency-like quantity balancing signaling speed against loss of response diversity. When optimized analytically, the framework predicts sparse scaling: the number of edges grows approximately linearly with the number of nodes. Sparsity is therefore not inserted as an engineering preference, it emerges from the trade-off. You can read more in a previous post:

When the same reasoning is integrated across time scales, hierarchical organization also appears, both in simulations and in comparisons with empirical networks.
Third, flows define a latent geometry. Entropy tells us how much response diversity the system has, but it does not by itself say where flows go. To answer that, one can build distances between nodes based on how they respond to perturbations over time.
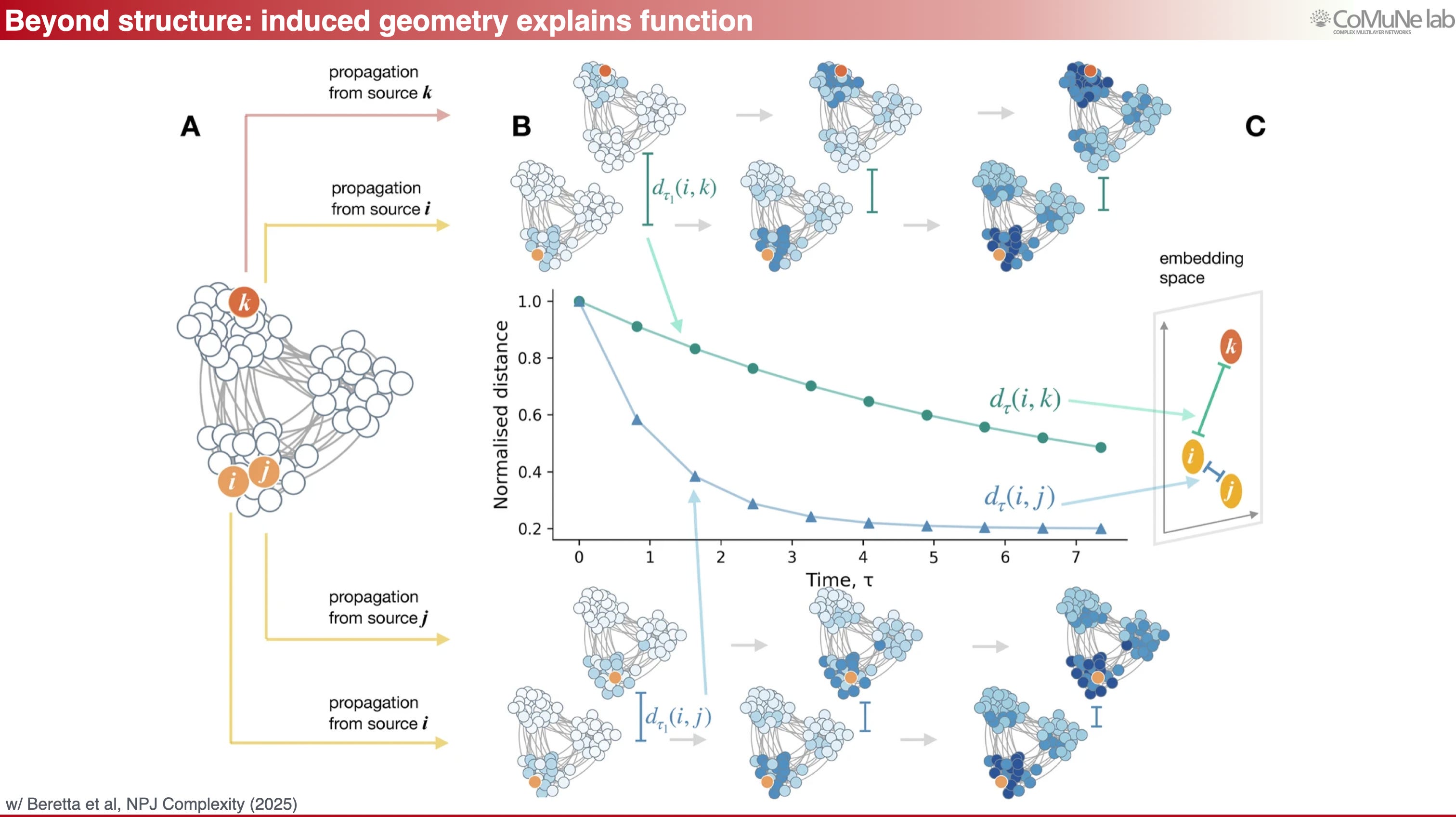
Nodes that exchange signals easily, or respond similarly, become close in latent space, while nodes that are functionally dissimilar become distant. In an application to brain networks, structural connectivity was combined with neuronal diffusion dynamics to construct such a distance matrix, and the resulting latent geometry was compared — and comparable — with a known mesoscale parcellation. The point is general: geometry here is not physical Euclidean geometry, it is a geometry induced by flows.
Take-home
The broader implication is that network science has to treat structure as a constraint, not as a complete explanation. Wiring matters since, without structure, flows have no substrate. But functionality lives in the propagation of perturbations, in the diversity of responses, in the persistence of flows across time scales and in the latent organization induced by dynamics.
This perspective is useful precisely because it travels across systems. Tardigrades, axolotls, connectomes, slime mold, fire ants, cities, transportation networks, metabolic pathways, and ecological simulations are not the same kind of object. They cannot — and should not — be forced into the same story at the level of substrate. They become comparable at the level of the phenomenon: how organized systems respond to perturbation, preserve diversity, coordinate flows and maintain accessible repertoires of behavior.
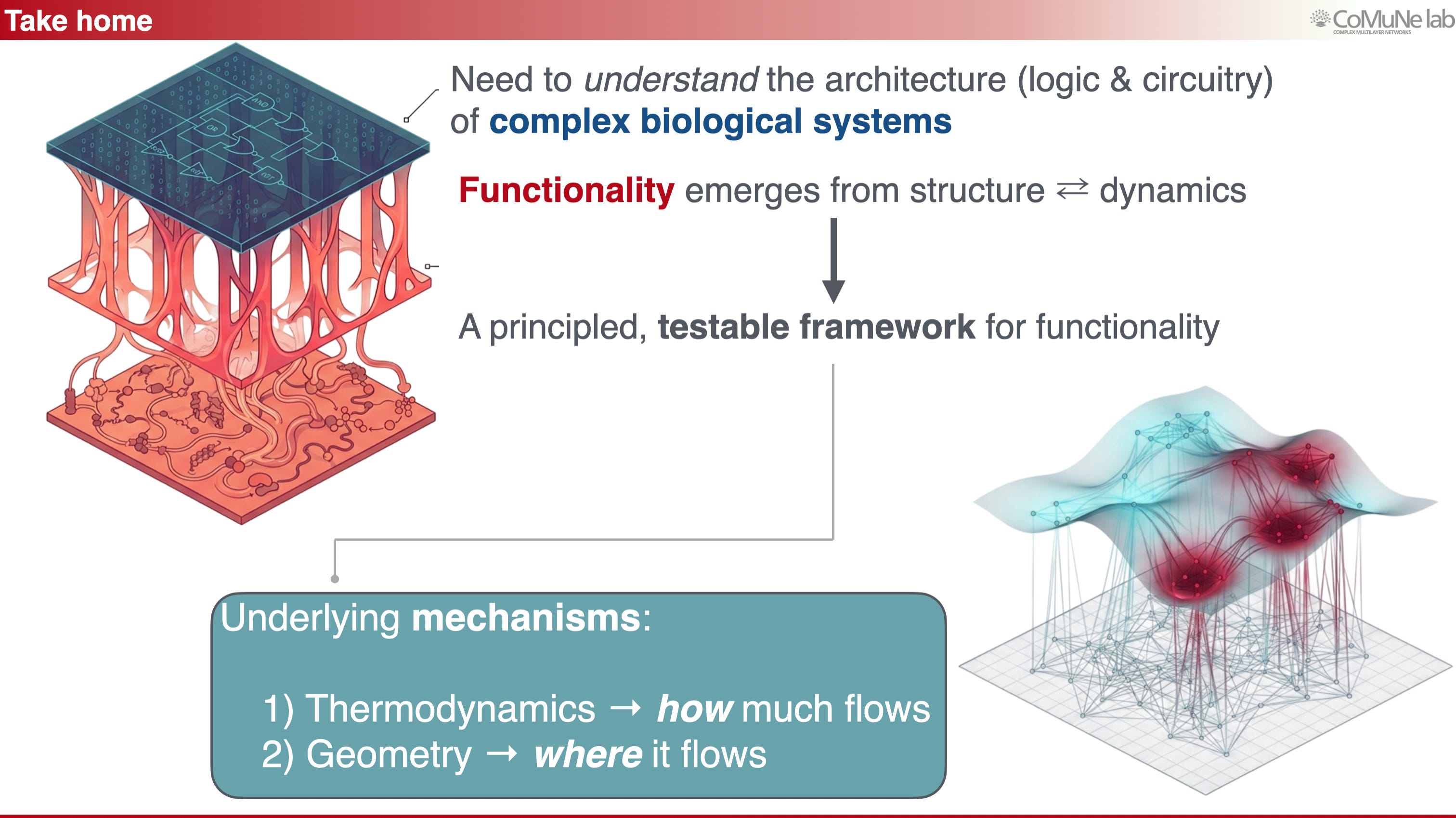
A theory of network functionality therefore needs two complementary tools:
Thermodynamics tells us how much flow a system can sustain under constraints.
Geometry tells us where that flow is organized.
Together, they move the analysis beyond the static map of connections: to understand what a complex adaptive system can do, we have to study how perturbations move through it.
→ If you still want to know more, maybe you should watch the video of the seminar.
→ Please, remind that if you find value in #ComplexityThoughts, you might consider helping it grow by subscribing, or by sharing it with friends, colleagues or on social media. See also this post to learn more about this space.